Trockenstressrisiko im Harz

Schneller Überblick
- Seit 1960 haben sich die Klimazonen im Harz verschoben
- Die Dauer extremer Trockenstressphasen ist dann in den Trockensommern 2018 bis 2020 etwa auf das Vierfache angestiegen
- Die Geschwindigkeit dieser Veränderungen überforderte offensichtlich die Anpassungsfähigkeit insbesondere der trockenheitsempfindlichen Fichten, wie das große Ausmaß der seit 2018 entstandenen Kalamitätsflächen verdeutlicht
Es ist unbestreitbar, dass die bei Wärme schnellere Entwicklung von Borkenkäfern hierzu einen wesentlichen Beitrag leistet. Unsere Untersuchung geht aber der Frage nach, welche Unterschiede es in der Dürreanfälligkeit der Baumarten gibt und wie diese schon ohne den Borkenkäfer im Harz vielleicht ein besonderes Risiko bei der Fichte begründen können. Dabei verwenden wir die im WKF-Verbundprojekt „Standortfaktor Wasserhaushalt im Klimawandel“ etablierte Methodik, um mit Wasserhaushaltssimulationen für Standardbestände die Auswirkungen unterschiedlicher Bodenbedingungen, Klimaverhältnisse und Baumarten räumlich hochaufgelöst für den gesamten Harz in Niedersachsen und Sachsen-Anhalt zu berechnen.
Boden und Klima
Grundlage für die Bodeninformation in der großflächigen Wasserhaushaltssimulation ist die forstliche Standortskartierung beider Bundesländer, durch die das gesamte Untersuchungsgebiet in über 62.000 Flächen mit jeweils einheitlichen Bodeneigenschaften eingeteilt ist. Den kartierten Lokalbodenformen im Harz Sachsen-Anhalts wurden Merkmalsspiegel (mittlere Bodeninformationen für die jeweilige Lokalbodenform) zugewiesen [1] (Tab. 1).
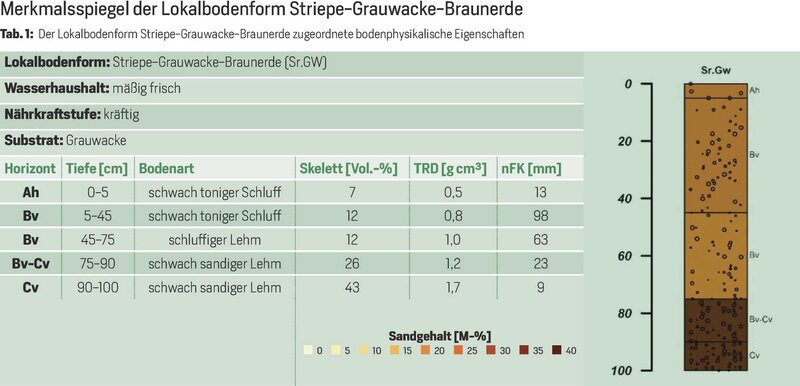
In Niedersachsen wurden den Standortstypen der Standortskartierung sog. Leitprofile zugeordnet. Hierzu wurden die verfügbaren Bodenprofile anhand des zugewiesenen forstlichen Standortstyps gruppiert und horizontbezogene Bodenparameter wie Bodenarten, Skelett- und Humusgehalte und Trockenrohdichten aggregiert [2]. Anhand der bodenphysikalischen Informationen können dann alle notwendigen Eingangsgrößen (hydraulische Leitfähigkeit, pflanzenverfügbares Bodenwasser usw.) geschätzt werden [3].
Die meteorologischen Größen für den gesamten Simulationszeitraum von 1961 bis 2020 wurden auf der Basis aller verfügbaren DWD-Stationen auf ein 250-m-x-250-m-Raster interpoliert (Weis et al., diese Ausgabe). Dabei wurden z. B. für die Niederschlagsinterpolation auf der Basis der bundesweit 4.752 Stationen Beziehungen zur Geländehöhe und zum beobachteten Zeitverlauf von Niederschlagsänderungen genutzt, um eine auch in bewegtem Gelände zutreffende Beschreibung der Niederschlagsverteilung zu erzeugen (Abb. 1). Die im Ergebnis vorliegende tägliche Meteorologie wurde auf die Flächen der forstlichen Standortskartierung übertragen.
Eigenschaften der Baumarten
An allen Standorten im Harz wurden gut wüchsige Standardbestände der Baumarten Buche, Eiche, Kiefer, Fichte und Douglasie miteinander verglichen. Die Baumartenunterschiede werden bei der Berechnung von Transpiration und Interzeption (Verdunstung vom Kronendach) durch die verschiedenen Eigenschaften der Baumarten wirksam (Tab. 2). So wurde die Belaubung jeder Baumart im Jahresverlauf auf Grundlage der maximalen Gesamtblattfläche im Sommer und der Zeitpunkte von Blattaustrieb und Laubfall klimaabhängig modelliert. Die verwendete Gesamtblattfläche des Kronendachs (Blattflächenindex, LAI) und die holzige Oberfläche (Holzoberflächenindex, SAI) beruhen dabei auf einem allometrischen Modell, das aus Daten von zahlreichen Waldbeständen abgeleitet wurde [4].
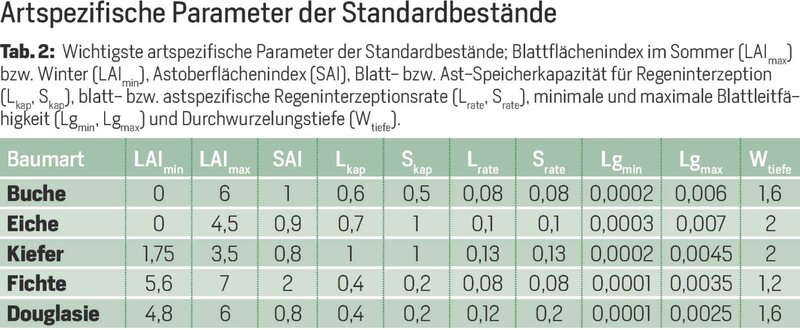
Zur Herleitung der Interzeptionsparameter (Speicherkapazitäten und blatt- bzw. astspezifische Interzeptionsraten) wurde die Wasserspeicherkapazität des Kronendachs, getrennt nach laublosem und belaubtem Zustand, für Niederschlagsereignisse von bis zu 37-jährigen Messreihen von 22 Intensivmonitoringflächen (262 Messjahre) bestimmt.
Die Transpiration der Baumarten wird neben der Blattfläche auch durch die Blattleitfähigkeit und die Wurzeltiefe gesteuert. Im Bereich der für die Baumgattungen veröffentlichten Wertespannen der Blattleitfähigkeit [5] wurden für die Wasserhaushaltsmodellierung der Monitoringflächen passende Werte durch Kalibrierung ermittelt. Die Durchwurzelungstiefe der Baumarten wurde in Anlehnung an Czajkowski et al. [6] festgelegt.
Veränderung des Klimas
Der über der norddeutschen Tiefebene weit herausgehobene Harz mit seinen starken Winden und häufigen Nebellagen nimmt eine klimatische Sonderstellung ein [7]. Wenn die auf den Wachstumsbedingungen für die Vegetation beruhende weltweite Klimazonen-Definition von Köppen/Geiger [8] auf die Waldgebiete im Harz angewendet wird, gehören für die Klimanormalperiode von 1961 bis 1990 die höchstgelegenen Kuppen des Harzes aufgrund der langen Phasen mit niedrigen Wintertemperaturen zur borealen Klimazone (Abb. 2). Das gilt außerhalb der Alpen wohl für kein anderes Gebiet in Deutschland.
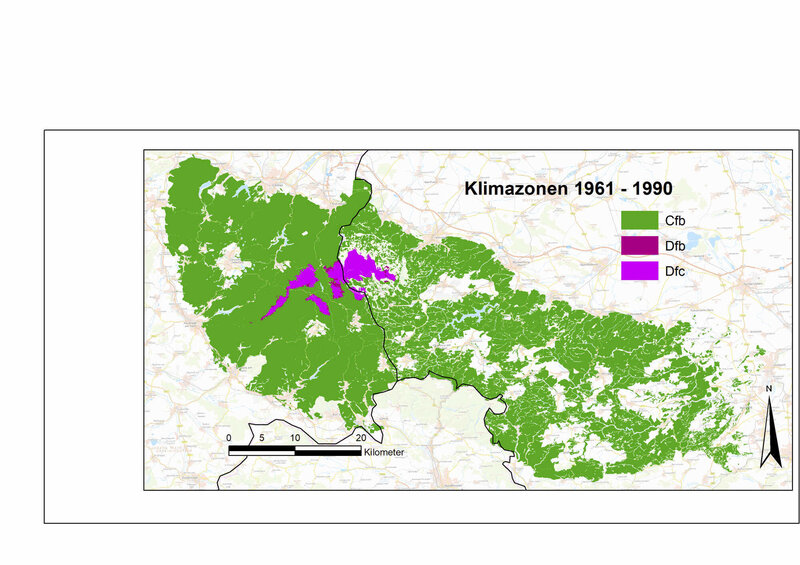
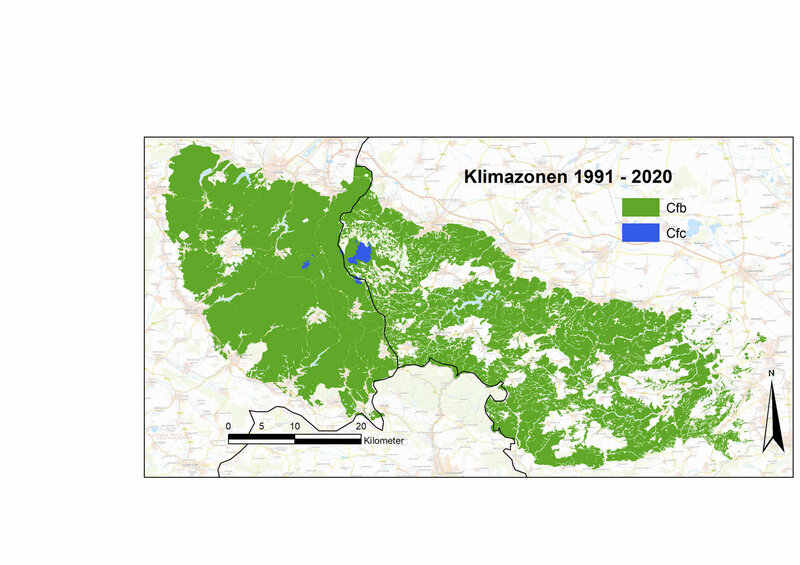
Bezogen auf die aktuelle Klimanormalperiode (1991 bis 2020) gibt es mittlerweile im Harz keine Areale der borealen Klimazone mehr. In 30 Jahren hat sich die Durchschnittstemperatur im Harz um ca. 1,0 K erhöht, gleichzeitig nahm der jährliche Niederschlag um ca. 40 mm zu, verringerte sich aber in der Vegetationsperiode um 7 mm. Als Folge des Klimawandels sind die Harzkuppen jetzt innerhalb der warm-gemäßigten Klimazone als humide Gebiete mit kühlen Sommern einzustufen, während der Rest des Harzes innerhalb derselben Klimazone zu den humiden Gebieten mit warmen Sommern gehört.
Veränderungen des Wasserhaushalts
Die unterschiedlichen Belaubungen und Blattleitfähigkeiten der Baumarten haben eine direkte Wirkung auf die Gesamtverdunstung von Wäldern, die sich aus Bodenevaporation, Interzeption und Transpiration zusammensetzt (Zahlenangaben im Folgenden immer als Durchschnitt der aktuellen Klimanormalperiode). Während Fichtenbestände mit ihrem hohen Blattflächenindex im Durchschnitt wenig Bodenevaporation zulassen (jährlich ca. 54 mm), weil der Boden durch das dichte Kronendach gut abgeschirmt ist, ist die Bodenevaporation bei den viel offeneren Kiefernbeständen generell am höchsten (98 mm). Im Vergleich der Bodenevaporation von 1961 bis 1990 mit der aktuellen Klimanormalperiode sind wenig Veränderungen zu erkennen, da die Zunahme des Verdunstungsanspruchs am Boden unter dem Kronendach noch gering ist.
Bei der Interzeption sind dagegen bei allen Baumarten überwiegend saisonale Veränderungen zwischen beiden Klimanormalperioden zu erkennen. Generell ist die Interzeption bei immergrünen Nadelbäumen und hier besonders bei Fichte und Douglasie am höchsten (449 mm bzw. 443 mm), während die laubabwerfenden Buchen und Eichen besonders im Winterhalbjahr sehr geringe Interzeptionsraten aufweisen (jährlich 231 mm bzw. 232 mm). In der Vegetationsperiode hat sich die Interzeptionsrate bei allen Baumarten um 5 bis 8 % erhöht, was hauptsächlich auf den Anstieg der Temperaturen zurückzuführen ist, durch die aufgefangenes Wasser schneller verdunstet. Gleichzeitig hat sich die Interzeptionsrate in der Vegetationsruhe im Winter aber um 6 bis 9 % verringert, da der Anteil der besonders effektiven Schneeinterzeption abgenommen hat. Aufs ganze Jahr gerechnet haben sich die Interzeptionsraten auf diese Weise kaum verändert.
Die Transpiration der Bäume ist ein Stoffwechselprozess, bei dem Wasser über die Wurzeln aus dem Boden aufgenommen und von den Blättern aufgrund des Wasserdampfdruckgefälles an die Atmosphäre abgegeben wird. Durch ihre Anpassung an frostbedingte Wassermangelsituationen im Winter ist die Blattleitfähigkeit der Nadelbaumarten generell geringer als bei den Laubbäumen, sodass auch die Transpiration niedriger ist (193 mm bis 244 mm bei Nadelbäumen, 265 mm bei Buche, 276 mm bei Eiche). Die höchste Transpirationsrate unter den drei Nadelbaumarten stammt dabei von Fichten: Weil bei ihnen die Gesamtblattfläche außerordentlich groß ist (vgl. Tab. 2), transpirieren sie auch mehr als z. B. Kiefern (227 mm), obwohl deren Blattleitfähigkeit je Blattfläche höher liegt (vgl. Tab. 2). Bei allen Baumarten gab es innerhalb von 30 Jahren eine Zunahme der Transpiration um 11 bis 13 %, die unmittelbar auf den temperaturbedingt höheren Verdunstungsanspruch der Atmosphäre zurückzuführen ist.
Dürreanfälligkeit der Baumarten
Das Spannungsverhältnis zwischen dem Verdunstungsanspruch der Atmosphäre (Tpot) und der tatsächlichen Wassernachlieferung durch die Bäume wird als Transpirationsquotient aus der Transpirationsrate T und der potenziellen Transpirationsrate (T/Tpot) ausgedrückt.
Der Transpirationsquotient resultiert somit aus dem tatsächlichen Zusammenwirken von Klima, Baumbestand und Bodenwasserverfügbarkeit. Bei nicht ausreichender Wasserversorgung oder bei Überforderung der Leitgefäße (z. B. bei trockener Luft und Wind) senken die Bäume die Wasserabgabe durch Schließen der Spaltöffnungen und der Index fällt unter 1 (= 100 %). Bei lang anhaltender Transpirationsreduktion (extreme Trockenstressphasen) reicht auch das permanente Schließen der Spaltöffnungen aber nicht mehr aus, weil eine minimale Transpiration weiter stattfindet. Die mit der Atmosphäre in Kontakt stehende wassergesättigte Oberfläche wird dann durch Einrollen der Blätter, Blatt- und Nadelverluste oder Leitgefäßembolien zusätzlich reduziert, mit oft bleibenden Schäden.
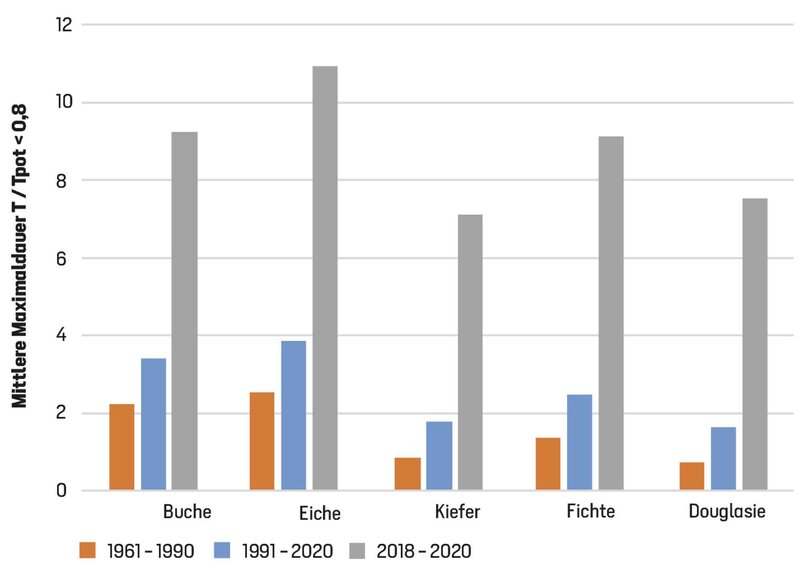
In Bezug auf solche Dürreschäden bei weitgehend geschlossenen Spaltöffnungen ist es deshalb interessant, wie lange extreme Trockenstressphasen andauern, in denen ein massives Absinken des Transpirationsquotienten auf z. B. Werte unter 80 % vorkommt. Die längste Phase mit derart extremem Trockenstress (Absinken von T/Tpot unter 80 %) dauerte im Durchschnitt der Klimanormalperiode 1961 bis 1990 im Harz bei den Laubbäumen zwischen 2,2 und 2,5 Tage, während sie bei den Nadelbäumen nur zwischen 0,7 Tage (Douglasie) und 1,4 Tage (Fichte) betrug (Abb. 3, vgl. Abb. 4 für die Unterschiede zwischen Standorten). Bei allen Baumarten hat sich die durchschnittliche Dauer der extremen Trockenstressphasen dann in der aktuellen Klimanormalperiode deutlich erhöht: Bei Buche und Eiche um jeweils 52 % auf 3,4 bzw. 3,9 Tage und bei den Nadelbäumen um 81 % bis 126 %. Die physiologische Notwendigkeit von adaptiven Wachstumsvorgängen (z. B. zur Verringerung des Verhältnisses von Gesamtblattfläche zur Wurzeloberfläche) war in diesem Zeitraum bei den Nadelbäumen etwa doppelt so groß wie bei den Laubbäumen. Unter den Nadelbäumen wies die Fichte die größte absolute Verlängerung der extremen Stressphasen auf, sie betrug hier im Durchschnitt +1,1 Tage, bei Douglasie und Kiefer verlängerten sich die extremen Stressphasen im Durchschnitt aller Standorte und Jahre um 0,9 Tage.
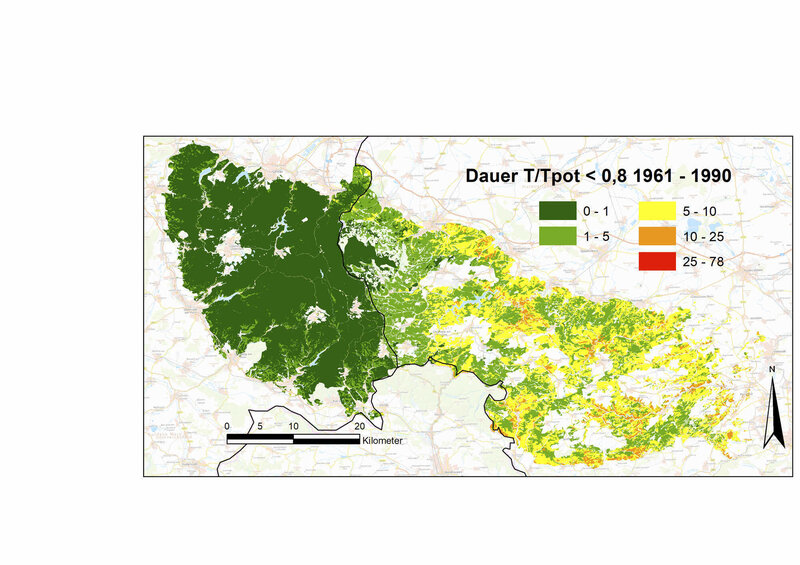
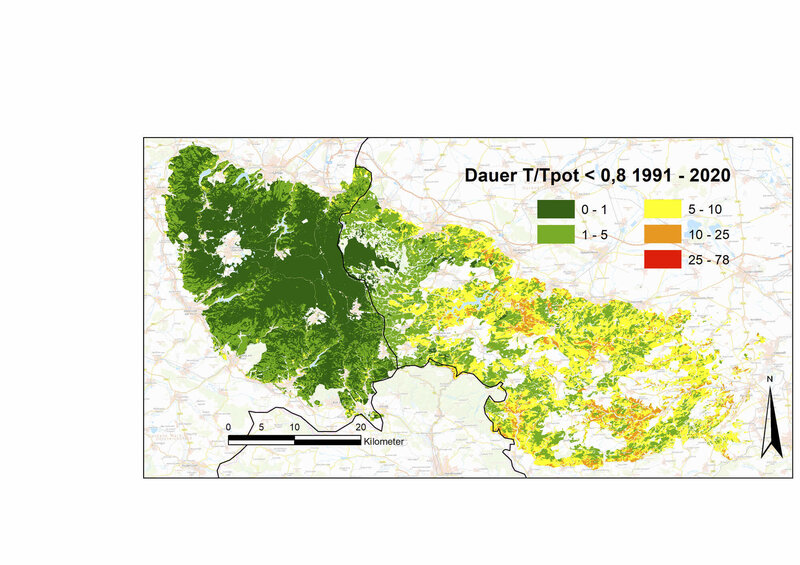
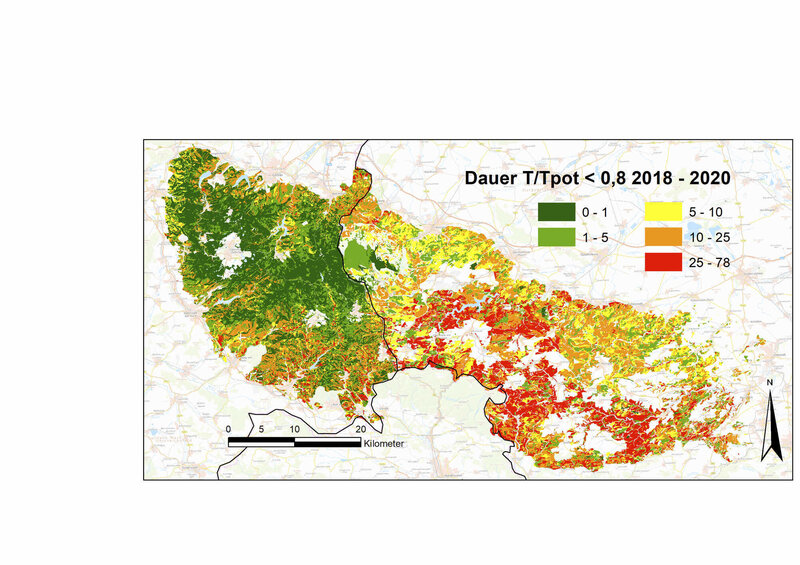
In den Jahren 2018 bis 2020 wurde dieser bei den Nadelbäumen schon vorher größere Anpassungsbedarf an lange und extreme Trockenstressphasen in drei aufeinanderfolgenden Jahren noch einmal drastisch gesteigert (Laubbäume +175 %, Nadelbäume +270 % bis +357 %, relativ zum Durchschnitt der Klimanormalperiode). Während Laubbäume außer mit bleibenden Schäden auch durch welke oder eingerollte Blätter reagieren können, können die Nadelbaumarten auf sehr lang anhaltenden extremen Trockenstress neben dem permanenten Spaltöffnungsschluss nur noch durch Nadelverlust oder/und Kavitation reagieren. Dass diese Phasen bei Fichten länger waren als bei Kiefern und Douglasien (durchschnittlich 9 Tage, an vielen Standorten über 25 Tage, Abb. 4) und somit die Schäden potenziell größer waren, lässt sich in dieser Berechnung auf ihre große Gesamtblatt- und Astoberfläche (große Oberflächen für Transpiration und Interzeption) bei gleichzeitig geringer Durchwurzelungstiefe und geringer maximaler Blattleitfähigkeit zurückführen.
„Der Klimawandel im Harz hat die Anpassungsfähigkeit von Fichtenbeständen überfordert.“
Fazit
Der Klimawandel führte im Harz zwischen 1990 und 2020 zu einer Verschiebung der Klimazonen. Die Dauer extremer Trockenstressphasen ist dann in den Dürresommern 2018 bis 2020 etwa auf das Vierfache angestiegen. Die Geschwindigkeit dieser Veränderung überforderte anscheinend die Anpassungsfähigkeit besonders der Fichten mit ihrer großen Gesamtblattfläche, weil sie die erforderliche Reduktion weiterer Wasserverluste nicht ohne massive Schäden erreichen können.
Literaturhinweise:
[1] AHRENDS, B.; HAFNER, S.; EVERS, J.; STEINICKE, C.; SCHMIDT, W.; MEESENBURG, H. (2016): Regionalisierung bodenphysikalischer Parameter für Waldstandorte in Sachsen-Anhalt. Beiträge aus der NW-FVA,14, 1–13. [2] HAFNER, S.; AHRENDS, B.; KÖHLER, B.; EVERS, J.; MEESENBURG, H. (2017): Ableitung von bodenphysikalischen Parametern für Waldstandorte in Schleswig-Holstein. Jahrestagung der DBG „Horizonte des Bodens“, 2.–7. September 2017. 1–8. [3] WESSOLEK, G.; KAUPENJOHANN, M.; RENGER, M. (2009): Bodenphysikalische Kennwerte und Berechnungsverfahren für die Praxis. Bodenökologie und Bodengenese, 40, 80 S. [4] WEIS, W.; AHRENDS, B.; BÖHNER, J.; FALK, W.; FLECK, S.; HABEL, R.; MEESENBURG, H.; PUHLMANN, H.; WEHBERG, J.; WELLPOTT A. (2021): Anhang zum Zwischenbericht 2020 des Projekts Standortfaktor Wasserhaushalt im Klimawandel (WHH-KW), FZK 22WK4141, Teilprojekte 01, 02, 03, 04; Projektförderung: Waldklimafonds (BMEL und BMU), 50 S. [5] HOSHIKA, Y.; OSADA, Y.; DE MARCO, A.; PENUELAS, J. (2018): Global diurnal and nocturnal parameters of stomatal conductance in woody plants and major crops. Glob. Ecol. Biogeogr., 27, 257–275. [6] CZAJKOWSKI, T.; AHRENDS, B.; BOLTE, A. (2009): Critical limits of soil water availability (CL-SWA) in forest trees – an approach based on plant water status. vTI agriculture and forest research 59(2), 87–93. [7] HERTEL, D.; SCHÖLING, D. (2011): Below-ground response of Norway spruce to climate conditions at Mt. Brocken (Germany) – A re-assessment of Central Europe’s northernmost treeline. Flora, 206, 127–135. [8] KOTTEK, M.; GRIESER, J.; BECK, C.; RUDOLF, B.; RUBEL, F. (2006): World Map of the Köppen-Geiger climate classification updated. Meteorol. Z., 15, 259–263; DOI: 10.1127/0941-2948/2006/0130.
Dr. Stefan Fleck
und Dr. Bernd Ahrends sind Mitarbeiter des Sachgebiets Intensives Umweltmonitoring in der Abteilung Umweltkontrolle der Nordwestdeutschen Forstlichen Versuchsanstalt in Göttingen. Dr. Henning Meesenburg ist Leiter des Sachgebiets.


✔ Immer und überall verfügbar – auf Ihrem Tablet, Smartphone oder Notebook
✔ Sogar im Offlinemodus und vor der gedruckten Ausgabe lesbar
✔ Such- und Archivfunktion, Merkliste und Nachtlesemodus